
Procariotele, care includ bacteriile și arheele, sunt organisme predominant unicelulare, adică nu prezintă nucleu și alte organite delimitate de membrane. Cromozomul bacterian are formă circulară și nu este organizat în jurul proteinelor denumite histone, ca în cazul cromozomilor de la eucariote. Regiunea centrală a celulei procariote în care se află ADN-ul poartă denumirea de nucleoid.
În afară de aceasta, procariotele conțin adesea numeroase plasmide, care sunt molecule de ADN scurte și circulare ce conțin una sau câteva gene. Plasmidele pot fi transmise în mod independent de cromozomul bacterian în timpul diviziunii celulare și adesea poartă trăsături precum cele implicate în rezistența la antibiotice.
Transcripția la procariote (la fel ca la eucariote) necesită ca dublul helix al ADN-ului să se desfășoare în regiunea de sinteză a ARN-ului mesager (ARNm). Acestă regiune poartă denumirea de buclă de transcripție. Transcripția începe întotdeauna de la aceeași catenă de ADN a fiecărei gene, care este denumită catenă matriță. Catena de ARNm rezultată este complementară catenei-matriță și este aproape identică cu cealaltă catenă de ADN (molecula de ADN este dublu catenară). Singura diferență este faptul că, în noua moleculă de ARNm, nucleotidele T (timină) sunt înlocuite cu nucleotide U (uracil). În catena dublu helix a ARN-ului, A (adenina) se poate lega de U prin intermediul a două legături de hidrogen, la fel cum A se leagă de T la nivelul ADN-ului.
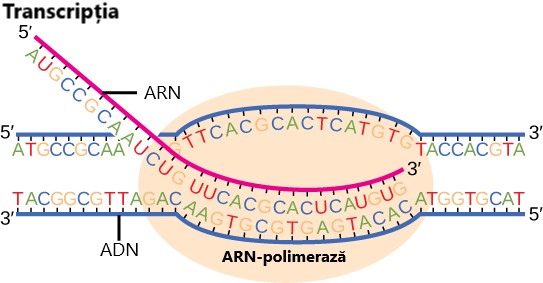
Molecula de ARN mesager este o copie de catenei de ADN de codare, cu diferența că T este înlocuită cu U. Sinteza ARN-ului are loc în direcția 5′ – 3′ și utilizează enzima ARN-polimerază.
Perechea de nucleotide din dublul helix ADN care corespunde situsului din care este transcrisă prima nucleotidă 5′ din ARNm se numește situs de inițiere. Nucleotidele care preced situsul de inițiere sunt notate cu „-”, iar cele care urmează situsului de inițiere sunt notate cu „+”.
Inițierea transcripției la procariote
Procariotele cu au un nucleu delimitat de o membrană. Drept urmare, procesele de transcripție, translație și degradare a ARNm au loc simultan. Nivelul intracelular de proteine bacteriene poate fi amplificat rapid prin multiple procese de transcripție și translație ce au loc la nivelul aceleași matrițe de ADN. Genomul procariotelor este foarte compact, iar transcripturile procariotelor acoperă adesea mai mult de o genă sau cistron (o secvență de codificare pentru o singură proteină). Moleculele de ARNm policistronic sunt apoi translatate pentru a produce mai mult de un tip de proteine.
ARN-polimeraza la procariote
Procariotele utilizează aceeași ARN-polimerază pentru transcripția tuturor genelor. La bacteria E. coli, polimeraza este alcătuită din cinci subunități polipeptidice, dintre care două sunt identice. Patru dintre aceste subunități, notate cu α, α, β și β’, reprezintă nucleul enzimei. Aceste subunități se asamblează de fiecare dată când o genă este transcrisă și se dezasamblează după finalizarea transcripției.
Fiecare subunitate îndeplinește un rol unic. Cele două subunități α sunt necesare pentru asamblarea polimerazei la nivelul ADN-ului, subunitatea β se leagă de ribonucleozid-trifosfatul ce va intra în structura moleculei de ARN, iar subunitatea β’ se leagă de catena-matriță de ADB. Cea de a cincea subunitate, notată cu σ, este implicată în inițierea transcripției. În lipsa sa, enzima ar desfășura procesul de transcripție în mod aleatoriu, producând molecule de ARNm la întâmplare. Polimeraza alcătuită din toate cele cinci subunități este numită holoenzimă.
Promotorii la procariote
Un promotor este o secvență de ADN la care mașinăria de transcripție, inclusiv ARN-polimeraza, se leagă și inițiază transcripția. În majoritatea cazurilor, promotorii sunt localizați înainte de de gena pe care o reglează. Secvența specifică a unui promotor este foarte importantă deoarece determină dacă gena corespunzătoare este transcrisă întotdeauna sau nu.
Deși promotorii variază la diferitele procariote, câteva elemente sunt comune la multe specii. La nivelul regiunilor -10 și -35 de dinaintea situsului de inițiere, există două secvențe consens sau regiuni care sunt similare la toți promotorii și la diferite specii bacteriene. Regiunea -10 are secvența consens TATAAT, iar regiunea -35 are secvența consens TTGACA. Aceste secvențe consens sunt recunoscute și legate de subunitatea σ. După realizarea aceste interacțiuni, subunitățile din nucleul enzimei se leagă de situs. Regiunea -10, bogată în A-T, facilitează desfășurarea matriței de ADN prin formarea mai multe legături fosfodiesterice. Faza de inițiere a transcripției se încheie cu producerea unor transcripturi de anulare, care sunt polimeri alcătuiți din aproximativ 10 nucleotide.
Faza de elongare
Faza de elongare începe cu eliberarea subunității σ a polimerazei, ceea ce permite enzime să continue de-a lungul matriței de ADN, sintetizând ARNm în direcție 5’ – 3’, cu o rată de aproximativ 40 de nucleotide pe secundă. Pe măsură ce elongarea continuă, ADN-ul este desfășurat înainte de nucleul și reînfășurat în urma acestuia. Împerecherea bazelor azotate în ADN și ARN nu este suficient de stabilă pentru a menține stabilitatea componentelor de sinteză a ARNm. În schimb, ARN-polimeraza acționează că un stabilizator între matrița de ADN și catena de ARN transcrisă pentru a se asigura că elongarea nu este întreruptă prematur.
Semnale de terminare
Odată ce o genă este transcrisă, polimeraza procariotelor trebuie să se disocieze de matrița de ADN și să elibereze noua catenă de ARNm. În funcție de gena care este transcrisă, există două tipuri de semnale de terminare: unul are la bază o proteină, iar celălalt are la bază ARN-ul.
Terminarea dependentă de rho este controlată de proteina rho (ρ), care urmează polimeraza de-a lungul lanțului de ARNm aflat în creștere. În apropiere de sfârșitul genei, polimeraza întâlnește un grup de nucleotide G pe matrița de ADN și se oprește. Drept urmare, proteina rho intră în coliziune cu polimeraze, ducând la eliberarea ARN-ului mesager din bucla de transcripție.
Terminarea independentă de rho este controlată de o secvență specifică de pe catena de ADN. Pe măsură ce polimeraza se apropie de sfârșitul genei transcrise, ea întâlnește o regiune bogată în nucleotide C-G. ARNm se pliază, iar nucleotidele complementare C-G se leagă. Rezultatul este un ac de păr stabil care face ca polimeraza să se blocheze imediată ce începe să transcrie o regiune bogată în nucleotide A-T. Regiunea U-A complementară de pe ARNm formează doar o legătură slabă cu matrița ADN, fapt care, coroborat cu blocarea polimerazei, induce suficientă instabilitate în nucleul enzimei pentru ca aceasta să elibereze noua moleculă de ARNm.
După terminare, procesul de transcripție este finalizat, ,iar noul ARNm procariotic format deja este utilizate pentru sinteze proteinelor codificate, deoarece ambele procese (transcripția și translația) pot avea loc simultan. Unificarea transcripției, translației și chiar a degradării ARN-ului mesager este posibilă pentru că toate aceste procese au loc în direcția 5’-3’ și pentru că celula procariotă nu prezintă compartimente delimitate de membrane. Prin contrast, prezența nucleului în celulele eucariote împiedică transcripția și translația simultane.