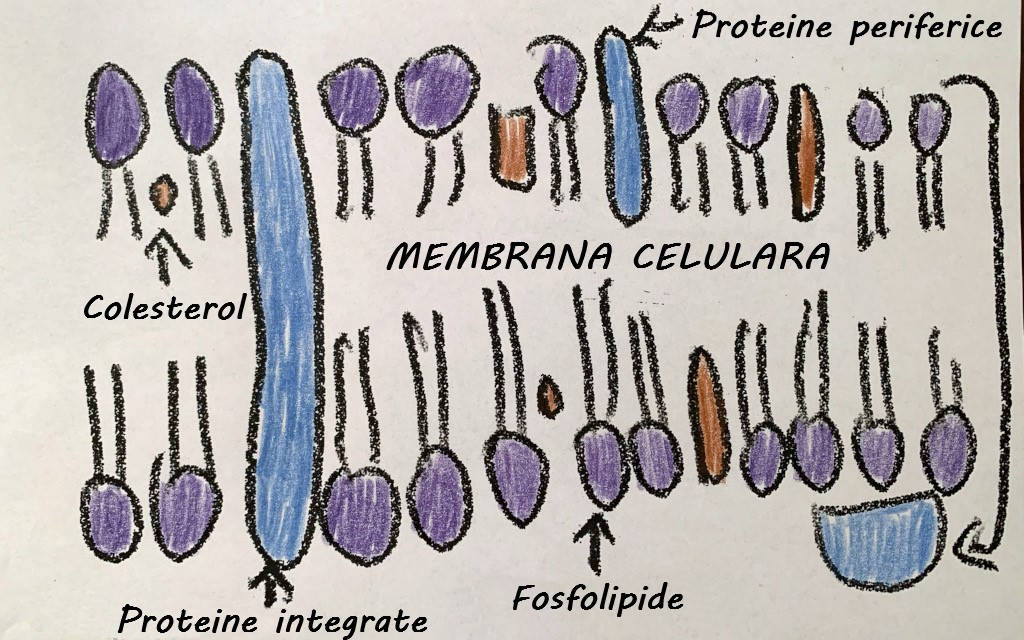
Membrana celulară (sau membrana plasmatică) este alcătuită din mai multe componente diferite. Fosfolipidele formează un strat dublu; cozile hidrofobe ale lor, compuse din acizi grași, sunt în contact unele cu altele, iar porțiunile hidrofile ale fosfolipidelor sunt orientate spre mediul apos intern și extern.
Alături de fosfolipide, în structura membranei celulare intră mai multe tipuri de proteine cu funcții diferite. Proteinele membranare integrate se întind adesea pe toată lățimea membranei și pot transporta materiale în sau din celule; aceste proteine încorporate pot fi hidrofile sau hidrofobe, în funcție de plasarea lor în membrană. Proteinele membranare periferice sunt întâlnite pe suprafețele exterioare și interioare ale membranelor și pot servi ca enzime, ca atașamente structurale pentru fibrele citoscheletului și ca parte a siturilor de recunoaștere ale unei celule. Aceste proteine „specifice celulei” joacă un rol vital în funcția imunitară, permit celulelor de un anumit tip (de exemplu, celule hepatice) să se identifice reciproc atunci când formează un țesut și permit hormonilor și altor molecule să recunoască celulele-țintă. Aceste proteine „plutesc” prin membrană, în flux constant.
Membrana plasmatică a unei celule definește celula, îi conturează granițele și determină natura interacțiunii acesteia cu mediul său. Celulele pot secreta, încorpora și excreta diferite substanțe, toate în cantități controlate. Membrana celulară trebuie să fie foarte flexibilă pentru a permite anumitor celule, cum sunt globulele roșii și globulele albe din sânge, să își schimbe forma pe măsură ce trec prin capilarele sanguine înguste.
Acestea sunt funcțiile mai importante ale unei membrane plasmatice. În plus, suprafața membranei plasmatice poartă markeri care permit celulelor să se recunoască între ele, ceea ce este vital pentru formarea țesuturilor și a organelor în timpul dezvoltării timpurii și care mai târziu joacă un rol în distincția „self” („propriu”) față de „non-self” („străin”) a răspunsului imun.
Printre cele mai sofisticate funcții ale membranei plasmatice se numără capacitatea de a transmite semnale prin intermediul unor proteine complexe, integrale, cunoscute sub numele de receptori. Aceste proteine acționează atât ca receptori ai intrărilor extracelulare, cât și ca activatori ai proceselor intracelulare. Acești receptori membranari furnizează locuri de atașare extracelulare pentru efectori, cum ar fi hormonii și factorii de creștere, și activează cascadele de răspuns intracelular atunci când efectorii lor sunt legați. Ocazional, receptorii sunt deturnați de virusuri care îi folosesc pentru a pătrunde în celule, iar, uneori, genele care codifică receptorii suferă mutații, determinând procesul de transducție a semnalului să funcționeze defectuos, ceea ce are consecințe dezastruoase.
Modelul mozaicului fluid
Existența membranei celulare a fost identificată în anii 1890, iar componentele sale chimice au fost identificate în 1915. Principalele componente identificate în acel moment au fost lipidele și proteinele.
Primul model acceptat pe larg al structurii membranei plasmatice a fost propus în 1935 de către savanții Hugh Davson și James Danielli; modelul se baza pe aspectul de „cale ferată” al membranei plasmatice în micrografiile electronice efectuate la acea vreme. Savanții au teoretizat că structura membranei plasmatice seamănă cu un sandviș, proteinele fiind analoage cu pâinea, iar lipidele fiind analoage cu umplutura.
În anii 1950, progresele în microscopie, în special microscopia electronică cu transmisie (TEM), au permis cercetătorilor să vadă că miezul membranei plasmatice constă dintr-un strat dublu, mai degrabă decât dintr-un singur strat. Un nou model care explica mai bine atât observațiile microscopice, cât și funcția membrane plasmatice a fost propus de S.J. Singer și Garth L. Nicolson în anul 1972.
Explicația propusă de Singer și Nicolson se numește modelul mozaicului fluid. Modelul a evoluat oarecum de-a lungul timpului, dar încă ține seama cel mai bine de structura și funcțiile membranei plasmatice așa cum le înțelegem acum.
Modelul mozaicului fluid descrie structura membranei plasmatice ca un mozaic de componente – inclusiv fosfolipide, colesterol, proteine și carbohidrați – care conferă membranei un caracter fluid. Membranele plasmatice variază de la 5 la 10 nanometri în grosime. Pentru comparație, globulele roșii umane, vizibile prin microscopie optică, au aproximativ 8 micrometri lățime, fiind de aproximativ 1.000 de ori mai late decât o membrană plasmatică. Membrana arată puțin ca un sandviș (Figura 1).

Figura 1 – Modelul mozaicului fluid al membranei celulare descrie membrana drept o combinație fluidă de fosfolipide, colesterol și proteine. Carbohidrații atașați de lipide (glicolipidele) și de proteine (glicoproteinele) se extind de pe suprafața exterioară a membranei.
Componentele principale ale unei membrane plasmatice sunt lipidele (fosfolipide și colesterol), proteinele și carbohidrații atașați la unele dintre lipide și unele dintre proteine. O fosfolipidă este o moleculă formată din glicerol, doi acizi grași și un grup principal legat de fosfat. Colesterolul, o altă lipidă compusă din patru inele de carbon fuzionate, se găsește alături de fosfolipidele din miezul membranei.
Proporțiile de proteine, lipide și carbohidrați din membrana plasmatică variază în funcție de tipul de celulă, dar pentru o celulă umană tipică, proteinele reprezintă aproximativ 50% din masă, lipidele (de toate tipurile) reprezintă aproximativ 40% din masă, restul de 10% din masă fiind carbohidrați. Cu toate acestea, concentrația de proteine și lipide variază în funcție de membranele celulare diferite. De exemplu, mielina, o componentă a membranei celulelor specializate care izolează axonii nervilor periferici, conține doar 18% proteine și 76% lipide. Membrana interioară mitocondrială conține 76% proteine și doar 24% lipide. Membrana celulară a globulelor roșii umane are 30% lipide. Carbohidrații sunt prezenți doar pe suprafața exterioară a membranei plasmatice și sunt atașați de proteine, formând glicoproteine, sau de lipide, formând glicolipide.
Fosfolipidele
Țesătura principală a membranei este compusă din molecule amfifile numite fosfolipide. Regiunile hidrofile (sau „iubitoare de apă”) ale acestor molecule sunt în contact cu fluidul apos atât din interiorul, cât și din exteriorul celulei. Moleculele hidrofobe (sau care „urăsc apa”) tind să fie nepolare. Ele interacționează cu alte molecule nepolare în reacții chimice, dar în general nu interacționează cu moleculele polare.
Atunci când sunt plasate în apă, moleculele hidrofobe tind să formeze o sferă sau un grup. Regiunile hidrofile ale fosfolipidelor tind să formeze legături de hidrogen cu apa și alte molecule polare atât pe exteriorul, cât și pe interiorul celulei. Astfel, suprafețele membranei care sunt orientate spre interiorul și exteriorul celulei sunt hidrofile. În schimb, interiorul membranei celulare este hidrofob și nu va interacționa cu apa. Prin urmare, fosfolipidele formează o excelentă membrană celulară cu două straturi, care separă fluidul din interiorul celulei de fluidul din exteriorul celulei.
O moleculă de fosfolipide (Figura 2) constă dintr-o coloană vertebrală de glicerol (o moleculă cu trei atomi de carbon), la care sunt atașate două molecule de acizi grași legate la atomii de carbon 1 și 2 și o grupare care conține fosfat atașată la al treilea carbon. Acest aranjament conferă moleculei o zonă denumită „cap” (grupul care conține fosfat), care are un caracter polar sau sarcină negativă, și o zonă denumită „coadă” (acizii grași), care nu are nicio sarcină. Capul poate forma legături de hidrogen, dar coada nu. O moleculă cu acest aranjament (o zonă încărcată pozitiv sau negativ și o zonă neîncărcată sau nepolară) este denumită amfifilă.

Figura 2 – O fosfolipidă este o moleculă cu doi acizi grași și o grupare fosfat modificată atașată la un schelet de glicerol. Fosfatul poate fi modificat prin adăugarea de grupări chimice încărcate sau polare.
Această caracteristică este vitală pentru structura unei membrane plasmatice, deoarece, în apă, fosfolipidele tind să se aranjeze cu cozile lor hidrofobe îndreptate una către cealaltă și capetele lor hidrofile îndreptate spre exterior. În acest fel, ele formează un dublu strat lipidic – o barieră compusă dintr-un strat dublu de fosfolipide care separă apa și alte materiale de pe o parte a barierei de apă și alte materiale de pe cealaltă parte. De fapt, fosfolipidele încălzite într-o soluție apoasă tind să formeze spontan mici sfere sau picături (numite micelii sau lipozomi), cu capetele lor hidrofile poziționate la exterior și cozile lor hidrofobe poziționate la interior.
Proteinele
Proteinele reprezintă a doua componentă majoră a membranelor celulare. Proteinele membranare integrate (unele tipuri specializate sunt numite integrine) sunt, după cum sugerează numele lor, integrate complet în structura membranei, iar regiunile lor hidrofobe interacționează cu regiunea hidrofobă a stratului dublu fosfolipidic.
Proteinele membranare integrate cu o singură trecere au, de obicei, un segment transmembranar hidrofob care constă din 20-25 de aminoacizi. Unele se întind doar pe o parte a membranei – asociindu-se cu un singur strat -, în timp ce altele se întind de pe o parte pe alta a membranei și sunt expuse pe ambele părți.
Unele proteine complexe sunt compuse din până la 12 segmente ale unei singure proteine, care sunt pliate extensiv și înglobate în membrană (Figura 3). Acest tip de proteină are o regiune sau regiuni hidrofile și una sau mai multe regiuni ușor hidrofobe. Acest aranjament tinde să orienteze proteina alături de fosfolipide, cu regiunea hidrofobă a proteinei adiacentă cozilor fosfolipidelor și regiunea sau regiunile hidrofile ale proteinei ieșind din membrană și intrând în contact cu citosolul sau lichidul extracelular.

Figura 3 – Proteinele cu membrane integrale pot avea unul sau mai multe alfa-helix-uri, care traversează membrana (exemplele 1 și 2) sau pot avea structuri beta-foaie-pliată ce se întind prin membrană (exemplul 3).
Proteinele membranare periferice se găsesc pe suprafețele exterioare și interioare ale membranelor, atașate fie de proteinele integrate, fie de fosfolipide. Proteinele periferice, împreună cu proteinele integrate, pot servi ca enzime, ca atașamente structurale pentru fibrele citoscheletului sau ca parte a siturilor de recunoaștere ale celulei. Acestea sunt uneori denumite proteine „specifice celulei”. Organismul își recunoaște propriile proteine și atacă proteinele străine asociate cu agenții patogeni invazivi.
Carbohidrații
Carbohidrații sunt a treia componentă majoră a membranelor plasmatice. Ei se găsesc întotdeauna pe suprafața exterioară a celulelor și sunt legați fie de proteine (formând glicoproteine), fie de lipide (formând glicolipide). Aceste lanțuri de carbohidrați pot consta din 2-60 de unități de monozaharide și pot fi liniare sau ramificate.
Alături de proteinele membranare periferice, carbohidrații formează situri specializate pe suprafața celulei care permit celulelor să se recunoască între ele. Aceste situri prezintă modele unice care permit recunoașterea celulei, la fel cum trăsăturile faciale unice pentru fiecare persoană îi permit să fie recunoscută. Această funcție de recunoaștere este foarte importantă pentru celule, deoarece permite sistemului imunitar să facă diferența între celulele corpului (numite „self”) și celulele sau țesuturile străine (numite „non-self”). Tipuri similare de glicoproteine și glicolipide se găsesc pe suprafețele virusurilor și se pot modifica frecvent, împiedicând celulele imune să le recunoască și să le atace.
Carbohidrați de pe suprafața exterioară a celulei – componentele carbohidraților atât ale glicoproteinelor, cât și ale glicolipidelor – sunt denumiți în mod colectiv drept glicocalix (însemnând „înveliș de zahăr”).
Glicocalixul este foarte hidrofil și atrage cantități mari de apă la suprafața celulei. Acest lucru ajută la interacțiunea celulei cu mediul său apos și la capacitatea celulei de a obține substanțe dizolvate în apă. Glicocalixul este, de asemenea, important pentru identificarea celulelor, pentru determinarea caracterului de „self” sau „non-self” și pentru dezvoltarea embrionară, fiind utilizat în atașamentele celulă-celulă pentru formarea țesuturilor.